
Чем археи отличаются от бактерий?
И бактерии, и археи – это микроорганизмы, обитающие в самых разных средах обитания, включая человеческое тело. Они очень похожи друг на друга даже под микроскопом. Однако их химический состав и физические характеристики сильно отличаются друг от друга. Вот некоторые из их ключевых отличий:
- Клеточные стенки и липиды мембран (жирные кислоты) бактерий и архей состоят из различных химических веществ;
- Многие виды бактерий могут выполнять фотосинтез (генерировать кислород из солнечного света), в то время как археи не могут;
- Жгутики архей и бактерий устроены по-разному;
- Археи размножаются путем деления, в то время как некоторые бактерии производят споры;
- Химический состав ДНК и РНК архей и бактерий сильно отличаются;
- Хотя некоторые бактерии являются патогенными (вызывают заболевание), археи не являются патогенными.
Особенности размножения и питания
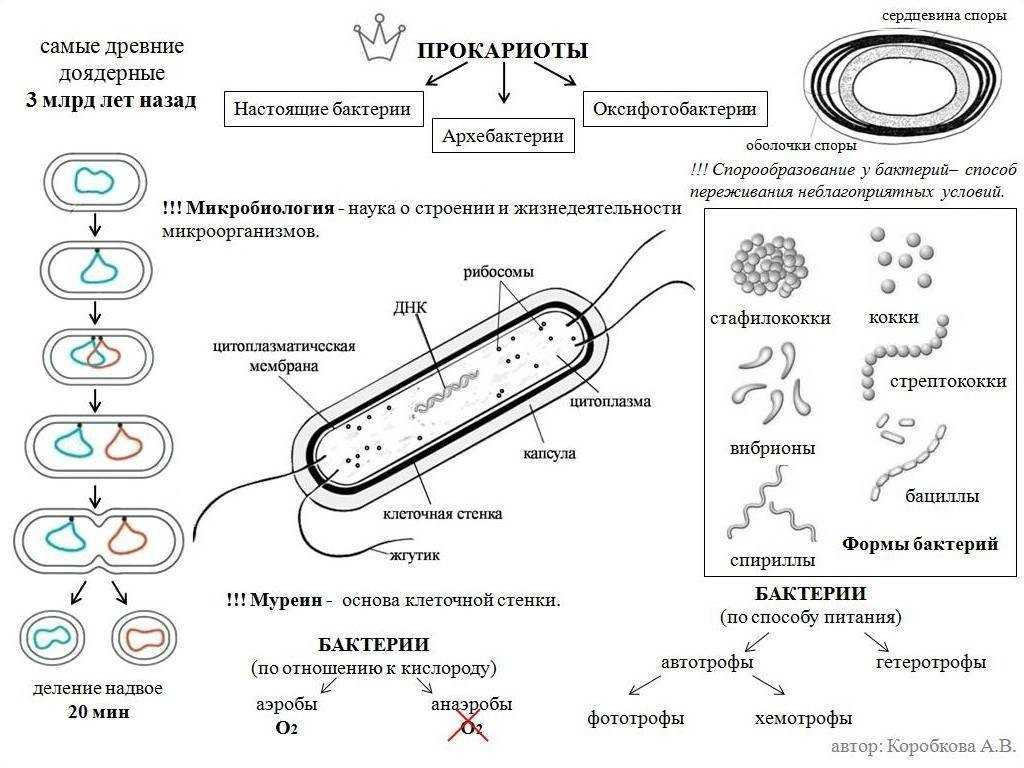
Особенности строения прокариотов таковы, что они размножаются только бесполым способом. Этот процесс осуществляется при попадании клеток в питательную среду, обеспечивающую активный рост.
Подавляющее большинство прокариотов размножается путём простого деления, которое происходит за счёт удвоения ДНК без образования хромосом. Две новые молекулы ДНК отделяются друг от друга плазматической мембраной, что приводит к делению клетки. В результате каждая дочерняя клетка содержит по одной молекуле ДНК.
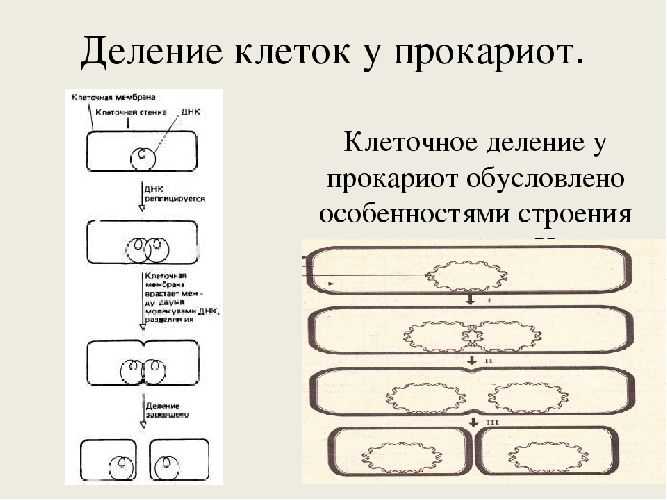 Рис. 2. Процесс деления прокариот.
Рис. 2. Процесс деления прокариот.
В благоприятных условиях процесс деления прокариот осуществляется каждые 25–30 минут. Этот интервал может увеличиться под воздействием сдерживающих факторов: дефицит пищи, солнечный свет, высокая температура и др.
По типу питания прокариоты бывают:
- Автотрофы — способные самостоятельно синтезировать органические вещества в процессе фотосинтеза (фототрофы) или получать энергию за счёт расщепления некоторых органических соединений (хемотрофы).
- Гетеротрофы — способные поглощать только готовые соединения.
Благодаря различным способам питания прокариоты смогли покорить разные среды обитания на планете.
Типы клеток и их развитие
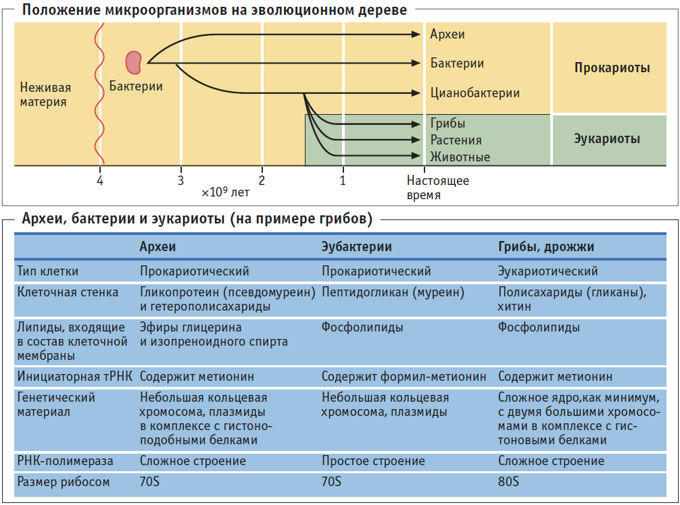
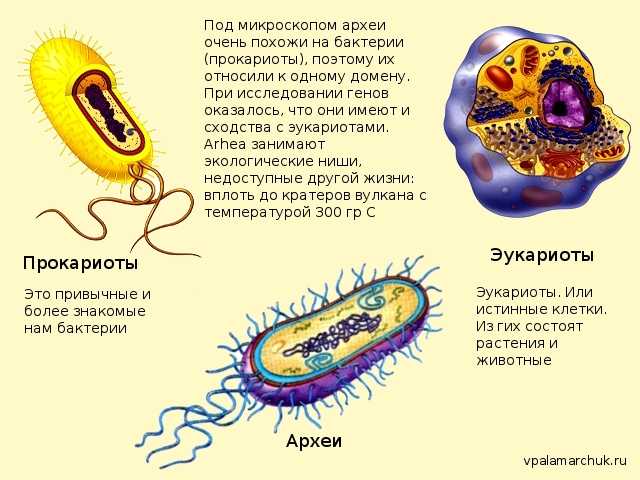
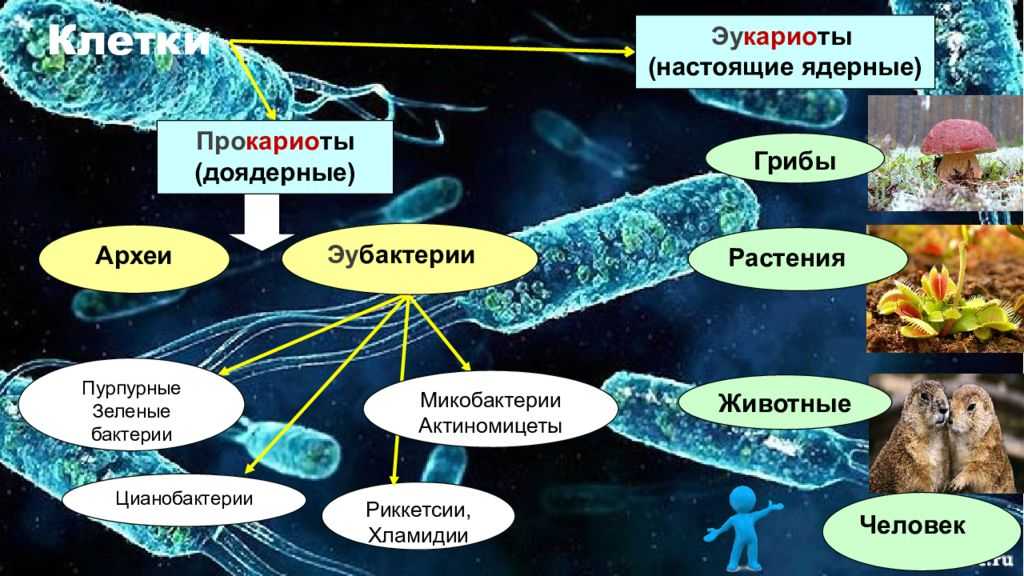
В течение многих лет считалось, что существует только два типа клеток: прокариоты (что буквально означает «без ядра») и эукариоты, более сложные и более поздние. Однако в последние два столетия они идентифицировали клетки другого типа, которые не соответствуют характеристикам любого из двух других.
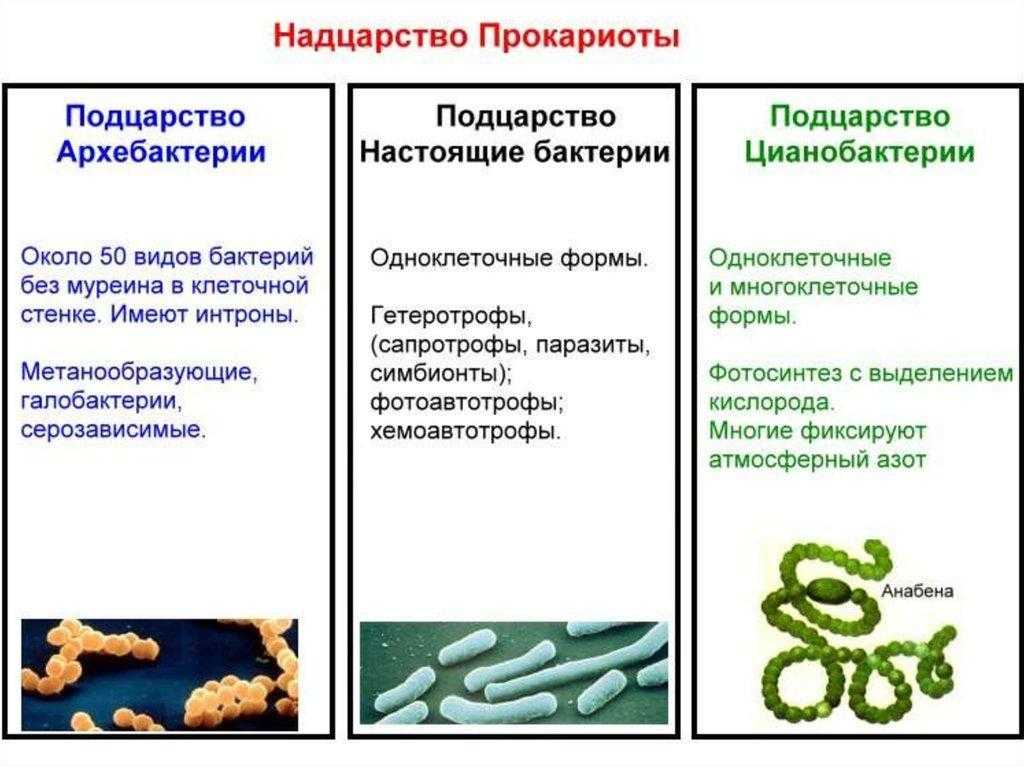
Эти клетки известны с 90-х годов как «археи», что буквально означает «старые». Таким образом, сегодня используется система классификации трех доменов: Archaea, Bacteria и Eucaria..
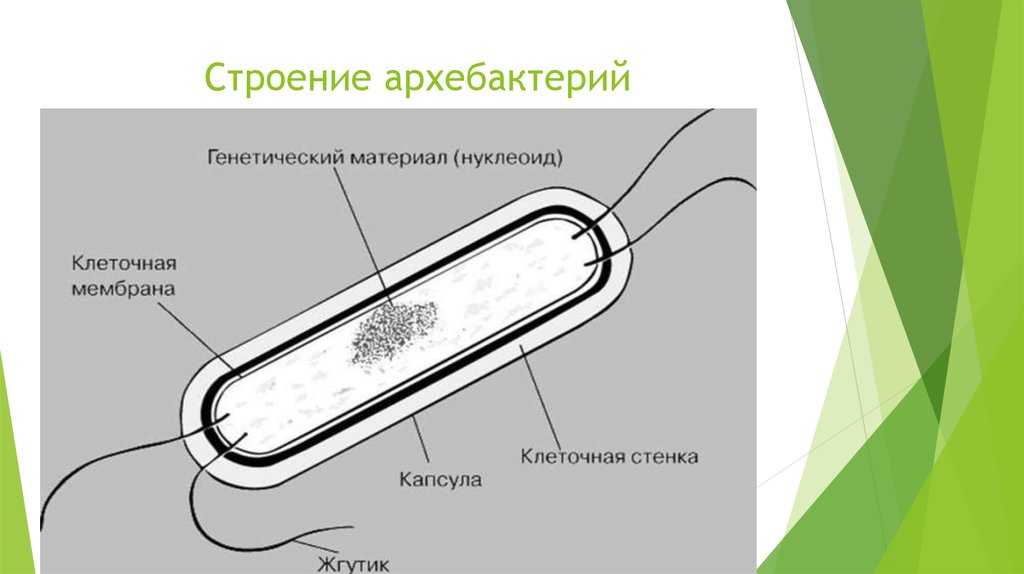
Клетки археи
Археи (также известные как Arqueas) — это клетки без ядра, очень похожие на бактерии, но с определенными характеристиками, которые заставляют их считаться независимыми организмами..
Как и остальные клетки, они являются микроскопическими организмами. Его клеточная стенка очень устойчива, что позволяет им жить в экстремальных условиях (даже в астероидах в космосе, без защиты какой-либо атмосферы).
Их диета также очень отличается, поскольку они используют неорганические соединения, такие как водород, диоксид углерода или сера вместо кислорода.
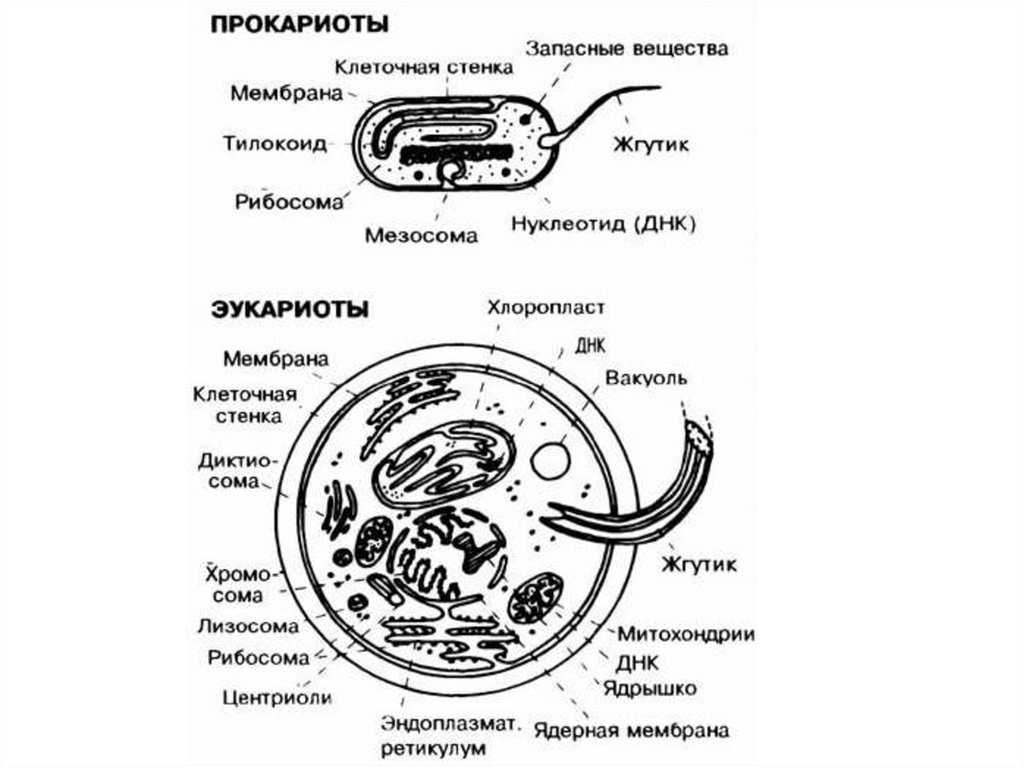
Прокариотические клетки (бактерии)
Прокариотические клетки являются простейшими среди трех типов. У них есть только клеточная мембрана, которая окружает внутреннюю часть клетки. Внутри мы можем найти генетический материал, взвешенный внутри цитоплазмы, а также некоторые рибосомы (органеллы, которые генерируют энергию внутри клетки).
Прокариотические клетки, несмотря на то, что они бывают разных типов, классифицируются как бактерии. Чтобы иметь возможность более эффективно адаптироваться к окружающей среде, многие из них имеют другие дополнения, такие как жгутики для свободного перемещения или липкая стенка, капсула, которая позволяет им прилипать к другим организмам..
Эукариотические клетки
Эукариотические клетки являются наиболее сложными и самыми крупными из трех типов. Они отличаются от прокариот и архей, в основном тем, что имеют ядро, в котором хранится ДНК. Кроме того, они имеют несколько типов клеточных органелл, которые позволяют им выполнять различные типы функций.
Эукариотические клетки являются основой всей сложной жизни, которая существует на Земле. Из-за этого ученые изучали их происхождение в течение многих десятилетий и разработали так называемую эндосимбиотическую теорию развития клеток..
Питание клеток прокариот
Питание — это процесс необходимый любому живому организму для получения энергии и веществ. Чтобы синтезировать органические соединения, живые организмы используют либо энергию света, либо энергию химических связей.
Организмы, способные к фотосинтезу, используют световую энергию. Они называются фототрофами. Для этого у них есть специфические особенности и способности.
Организмы, использующие только химическую энергию, называются хемотрофами.
Большая часть организмов относится к гетеротрофам, т.е. к организмам, которые используют готовые органические вещества.
Прокариоты, по способу питания делятся на две группы:
• Автотрофы (синтезируют органические вещества из неорганических):
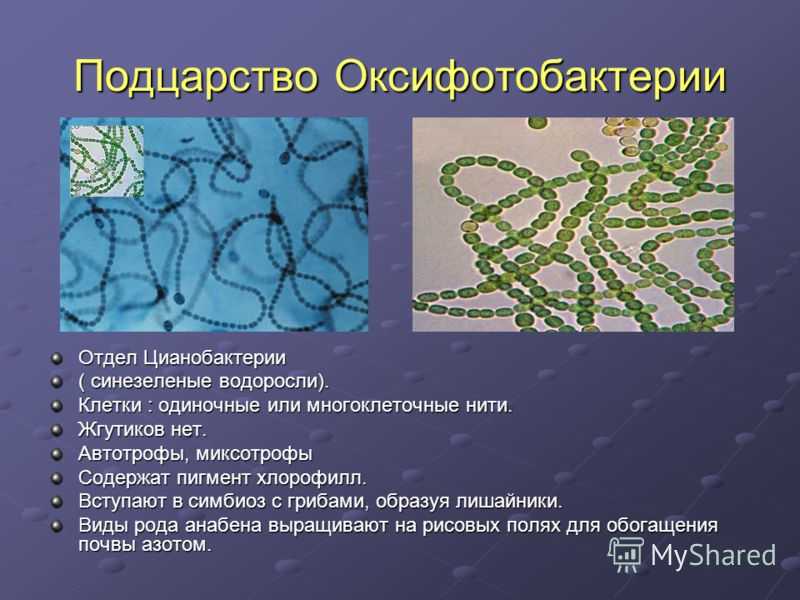
o фотосинтезирующие сине-зеленые или цианобактерии. За счет способности к фотосинтезу, они выделяют кислород. Некоторые из них могут связывать газообразный азот воздуха, и переводить его в состав азотсодержащих органических веществ;
o хемосинтезирующие (железобактерии и нитрифицирующие бактерии) получают энергию в процессе окисление неорганических веществ таких как аммиак и нитраты.
• Гетеротрофы (используют готовые органические вещества):
o сапротрофы – питаются мертвыми органическими веществами. Бактерии выделяют на мертвую органику ферменты, и переваривание пищи у них происходит вне организма. Уже растворимые продукты поступают в тело сапротрофа (бактерии гниения и брожения);
o симбионты (явление мутуализма) – организмы, живущие в симбиозе с другими организмами и получающие органические вещества от них (клубеньковые бактерии, бактерии кишечника человека, которая обеспечивает человеческий организм витаминами групп В и К);
o паразиты питаются органическими веществами живых организмов (хозяев), внутри которых они обитают (болезнетворные бактерии или микробы). Бактерии, вызывающие болезни называются патогенами. Паразиты, которые могут жить только в живых клетках, называются облигатными. Но есть бактерии, которые сначала заражают «хозяина» и вызывают его гибель, а потом живут как сапротрофы.
Аэробы – используют для дыхания атмосферный кислород (бактерии гниения); анаэробы живут в отсутствии кислорода (бактерии ботулизма).
Эндосимбиотическая теория эволюции клеток
Эукариотические клетки гораздо более развиты, чем археи или бактерии. Всего несколько десятилетий назад было найдено удовлетворительное объяснение его возникновения: теория эндосимбиотика.
Эта теория основана на сходстве между митохондриями и хлоропластами эукариотических клеток с бактериями, как по форме, так и по функционированию..
Поэтому ученые, защищающие его, предполагают, что в какой-то момент эволюции большая клетка впитала бактерию и начала использовать ее для извлечения энергии, необходимой для выживания и размножения..
Поглощенные бактерии, с другой стороны, получили большую вероятность оставить потомство, а также большую безопасность, находясь внутри более крупной клетки. Поэтому были симбиотические отношения; отсюда и название теории.
После миллионов лет эволюции, митохондрии и хлоропласты, которые раньше были независимыми бактериями, стали специализированными. Поэтому они больше не могут выживать вне клетки.


Доказательства эндосимбиотической теории
В повседневном языке мы используем слово «теория», чтобы описать мнение, которое не основано на фактах. Однако в мире науки теория — это объяснение явления, подтвержденного экспериментами и наблюдениями..
Эндосимбиотическая теория не является исключением. Несколько подсказок заставляют нас думать, что именно так появились клетки животных и растений. Вот некоторые из этих доказательств:
- Митохондрии и хлоропласты имеют свою собственную ДНК → Эти два типа органелл являются единственными, которые имеют ДНК внутри своей цитоплазмы, отделенные от основной ДНК клетки.
- Обе органеллы размножаются самостоятельно → Поскольку они имеют свою собственную ДНК, хлоропласты и митохондрии могут реплицироваться независимо от клетки и направлять свое собственное деление.
- У них есть клеточная мембрана → В отличие от остальных органелл в клетке, и митохондрии, и хлоропласты имеют двойную клеточную мембрану, которая отделяет их от остальных. Этот тип мембраны также присутствует в бактериях.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.
Размножение
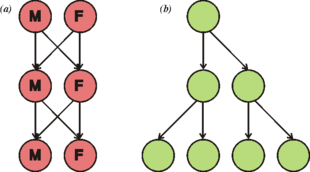
Деление клеток обычно происходит бесполым путем посредством митоза, процесса, который позволяет каждому дочернему ядру получить по одной копии каждой хромосомы. У большинства эукариот также есть жизненный цикл, который включает половое размножение, чередование между гаплоидной фазой, когда только одна копия каждой хромосомы присутствует в каждой клетке и диплоидная фаза, в которой две копиикаждой хромосомы присутствуют в каждой клетке. Диплоидная фаза образуется в результате слияния двух гаплоидных гамет с образованием зиготы, которая может делиться митозом или подвергаться хромосомной редукции в результате мейоза. Этот паттерн сильно различается. У животных нет многоклеточной гаплоидной фазы, но каждое поколение растений может состоять из гаплоидной и диплоидной многоклеточных фаз.
Эукариоты имеют меньшее отношение площади поверхности к объему, чем прокариоты,и, следовательно, имеют более низкую скорость метаболизма и более длительное время генерации.
Эволюция полового размножения может быть изначальной и фундаментальная характеристика эукариот. Основываясь на филогенетическом анализе, Дакс и Роджер предположили, что факультативный пол присутствовал у общего предка всех эукариот. Основной набор генов, которые функционируют в мейозе, присутствует как в Trichomonas vaginalis, так и в Giardia Кишечник, двухорганизмах, которые ранее считались бесполыми. Поскольку эти два вида являются потомками линий, которые рано разошлись с эволюционным деревом эукариот, был сделан вывод, что основные мейотические гены и, следовательно, пол, вероятно, присутствовали у общего предка всех эукариот. Было показано, что виды эукариот, которые когда-то считались бесполыми, например паразитические простейшие из рода Leishmania, имеют половой цикл. Кроме того, теперь данные показывают, что амебы, ранеесчитавшиеся бесполыми, в древности являются половыми и что большинство современных бесполых групп, вероятно, возникли недавно и независимо.
7) Первые рептилии. Синапсиды.
Начавшийся примерно 360 млн. лет назад и продолжавшийся 60 млн. лет каменноугольный период был очень благоприятен для земноводных. Значительную часть суши покрывали болота, климат был тёплым и влажным. В таких условиях многие земноводные продолжали жить в воде или около неё. Но примерно 340-330 млн. лет назад некоторые из земноводных решили освоить и более сухие места. У них развились более сильные конечности, появились более развитые лёгкие, кожа, наоборот стала сухой, чтобы не терять влагу
Но чтобы действительно длительное время жить далеко от воды, нужно было ещё одно важное изменение, ведь земноводные, как и рыбы, метали икру, и их потомство должно было развиваться в водной среде. И около 330 млн
лет назад появились первые амниоты, т. е. животные, способные откладывать яйца. Оболочка первых яиц была ещё мягкой, а не твёрдой, тем не менее, их уже можно было откладывать на суше, а значит, потомство уже могло появляться вне водоёма, минуя стадию головастиков.
Учёные до сих пор путаются в классификации земноводных каменноугольного периода, а также в том, считать ли некоторые ископаемые виды уже ранними рептилиями, либо всё ещё земноводными, приобретшими лишь некоторые черты рептилий. Так или иначе, эти то ли первые рептилии, то ли рептилоподобные земноводные выглядели примерно так:

Вестлотиана — небольшое животное длиной около 20 см., сочетавшее черты рептилий и земноводных. Жило примерно 338 млн. лет назад.
А затем ранние рептилии разделились, дав начало трём большим группам животных. Палеонтологи выделяют эти группы по строению черепа — по числу отверстий, через которые могут проходить мышцы. На рисунке сверху вниз черепа анапсида, синапсида и диапсида:
При этом анапсидов и диапсидов часто объединяют в группу завропсидов. Казалось бы, отличие совершенно незначительное, тем не менее, дальнейшая эволюция этих групп пошла совершенно разными путями.
От завропсидов произошли более продвинутые рептилии, включая динозавров, а затем птицы. Синапсиды же дали начало ветви звероподобных ящеров, а затем и млекопитающим.
300 млн. лет назад начался Пермский период. Климат стал более сухим и холодным и на суше стали доминировать ранние синапсиды — пеликозавры. Одним из пеликозавров был Диметродон, имевший в длину до 4х метров. На спине у него был большой «парус», который помогал регулировать температуру тела: быстро охладиться при перегреве или, наоборот, быстро согреться, подставив спину солнцу.

Считается, что огромный диметродон является предком всех млекопитающих, а значит, и человека.
Происхождение эукариот и половое размножение
Появление эукариот — второе по значимости событие в истории земной жизни (первое — само появление жизни). Эукариотическая клетка устроена гораздо сложнее, чем прокариотическая, а промежуточные формы между ними, существовавшие когда-то, давно вымерли. Поэтому вопрос о происхождении эукариот остается одной из самых сложных и спорных тем в эволюционной биологии (см.: А. В. Марков, А. М. Куликов, 2009. Происхождение эукариот как результат интеграционных процессов в микробном сообществе). Правда, недавно ситуация резко изменилась к лучшему в результате открытия локиархей — неизвестной ранее группы архей, обладающей многими признаками, которые до сих пор считались уникальными для эукариот.
Но даже с учетом локиархей получается, что эволюционный путь от последнего общего предка с ближайшими архейными родственниками до последнего общего предка всех современных эукариот (LECA, last eukaryotic common ancestor) был долгим и трудным. На этом пути предки эукариот приобрели целый ряд признаков, которых нет ни у кого из прокариот, даже у локиархей. Одним из последних шагов на этом пути, по-видимому, стало приобретение внутриклеточных симбионтов — альфапротеобактерий, давших начало митохондриям.
К числу эукариотических инноваций, наиболее трудных для объяснения, относится эукариотический секс (называемый также амфимиксисом или, попросту, половым размножением). Это специфический и очень эффективный способ перемешивания генетического материала разных особей, включающий сингамию (слияние гаплоидных клеток или ядер, приводящее к удвоению хромосомного набора) и мейоз (особый вариант клеточного деления, приводящий к сокращению числа хромосом вдвое). Мейоз сопровождается кроссинговером, в ходе которого гомологичные хромосомы обмениваются гомологичными участками.
Прокариотический секс устроен гораздо проще: это однонаправленная передача небольших фрагментов генетического материала от микроба-донора микробу-реципиенту (см. врезку и рис. 2).
Развитие первых клеток
Первые ячейки появились по крайней мере 3700 миллионов лет назад, примерно через 750 миллионов лет после образования Земли. Хотя мы не знаем наверняка, как появились первые клетки, мы достаточно хорошо знаем, как они развивались.


![Глава 5 ранний докембрий: древнейшие следы жизни на земле. маты и строматолиты. прокариотный мир и возникновение эукариотности. удивительная палеонтология [история земли и жизни на ней]](https://zd-semya.ru/wp-content/uploads/e/f/4/ef4ebf1c3ff09f325dad5709478a6650.jpeg)
Тем не менее, одна из наиболее принятых теорий о формировании первых ячеек заключается в следующем: учитывая атмосферные условия примитивной Земли, разряд энергии может производить органические молекулы, чтобы спонтанно образоваться.
Это было продемонстрировано экспериментами Стэнли Миллера в 50-х годах, в которых ему удалось создать органические молекулы из водорода, метана и аммиака..
Позже были сформированы первые сложные органические молекулы (также называемые макромолекулами). В какой-то момент эволюции этих молекул первая была способна копировать, используя материалы из окружающей среды. Он родился тогда, впервые в камере.
Эти первые клетки могли сначала размножаться свободно, учитывая отсутствие конкуренции за топливо, которое они использовали. Однако, поскольку их количество значительно увеличилось (именно из-за отсутствия конкуренции), вскоре клетки должны были стать более изощренными, чтобы продолжить размножение. Так начался процесс эволюции.
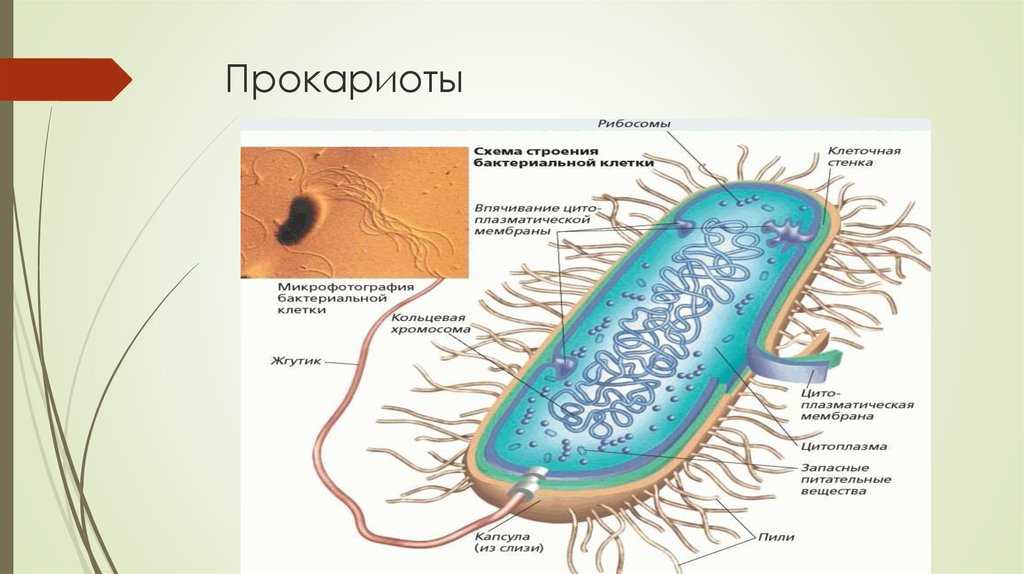
Особенности клеток прокариот
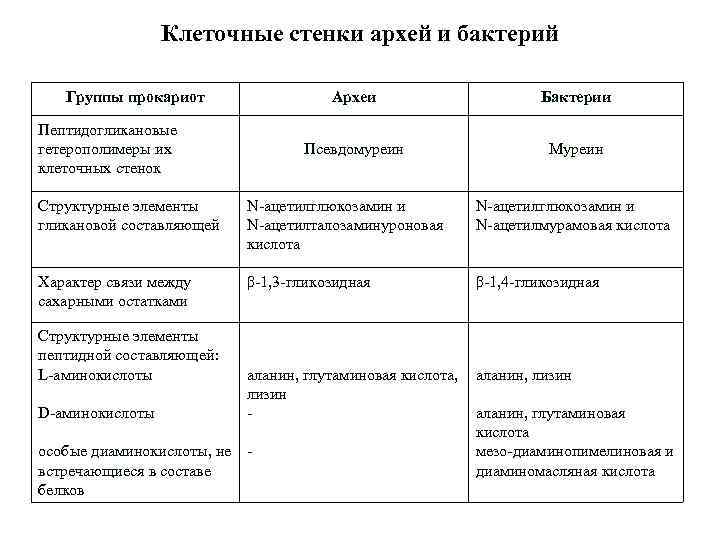
Клеточная стенка прокариот состоит из белка, который называется муреин и слизистой капсулы из полисахаридов. Внутри клетки располагается нуклеоид – это бактериальная хромосома, которая представляет собой одиночную кольцевую молекулу ДНК.
Также в клетках располагаются рибосомы, с помощью которых происходит синтез белка. У фотосинтезирующих бактерий в клетку встроены фотосинтетические мембраны (о них будет сказано дальше) и органелла дыхания или мезосома.
Оболочка клеток прокариот могут иметь выросты – жгутики и пили. С их помощью клетка передвигается. Прокариоты не содержат митохондрий (своеобразная энергетическая станция). Синтез АТФ (источник энергии для всех биохимических процессов в живых клетках) происходит на плазматической мембране.
Обратите внимание! У клеток есть органеллы – это специализированные микроструктуры, которые постоянно есть в клетках и которые выполняют ряд жизненно важных функций (внутриклеточный обмен веществ, энергетический и информационный обмен)
Гипотезы эндосимбиотического происхождения других органелл
В то время как эндосимбиоз является наиболее принятой версией происхождения митохондрий и пластид, попытки применения теории симбиогенеза к другим органеллам и структурам клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предпололожить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи.
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза (англ. viral eukaryogenesis). В ее основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра.
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующший аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах противоречивы. однако большинство исследователей ее наличие отрицают. Увеличение числа базальных телец и гомологичных им центриолей происходит не путем деления, а путем достраивания нового органоида рядом со старым.
Сходства и различия в молекулярных процессах, протекающих в клетках прокариот и эукариот
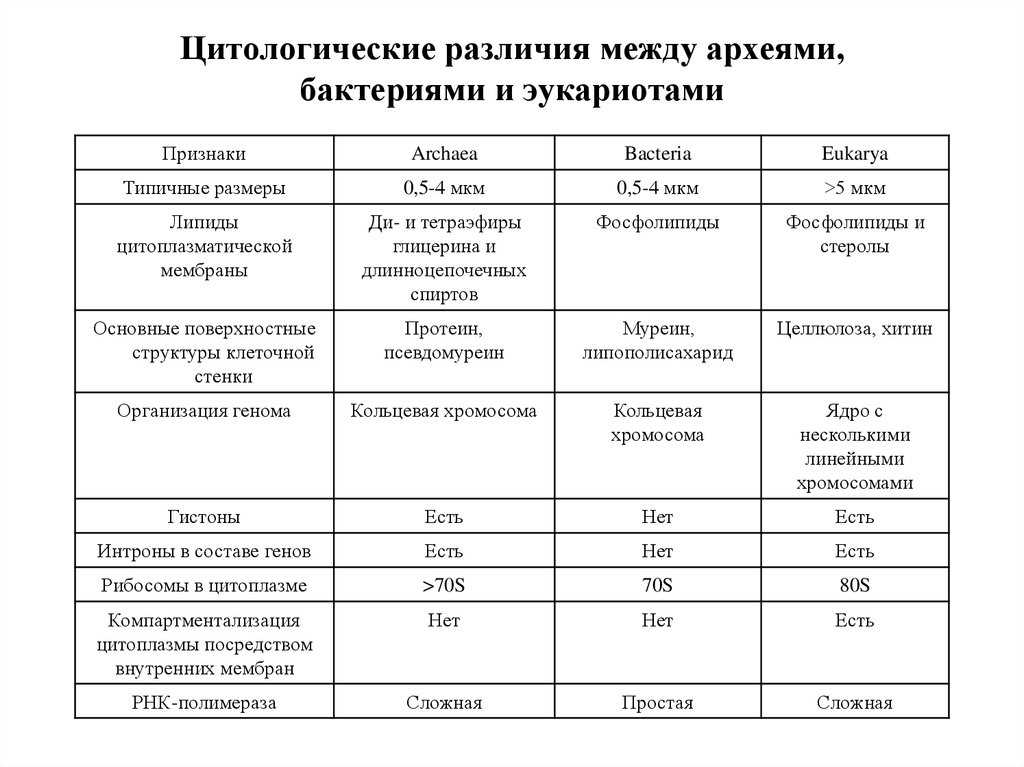
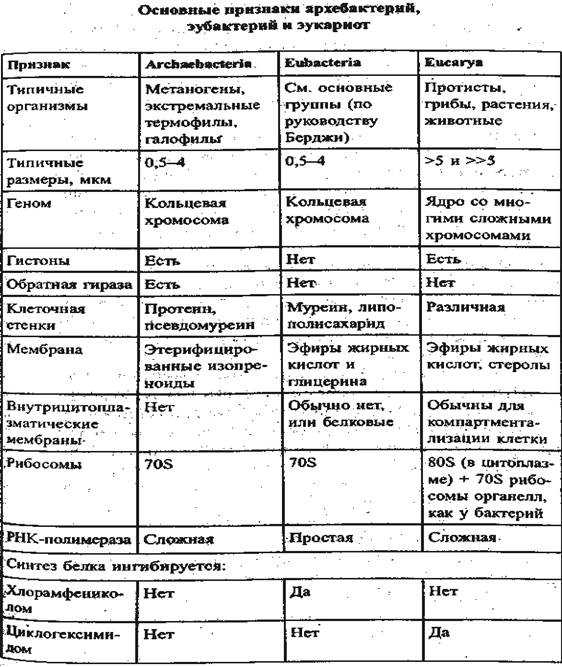
Различия в организации генетического материала для этих групп не ограничиваются лишь его расположением и тем, замкнута ли ДНК в кольцо. Процессы транскрипции и трансляции у каждой группы имеют свои особенности. Например, для поддержания структуры ДНК и регуляции экспрессии генов в клетках эукариот и архей есть специальные белки – гистоны, которых нет у бактерий.
Гены бактерий собраны в опероны. Это означает, что несколько генов находятся друг за другом и имеют общий промотор (место старта трансляции), таким образом мРНК получается полицистронная, то есть кодирующая несколько белков. Эта особенность характерна и для архей. У эукариот, наоборот, для каждого гена есть свой промотор. В то же время, общим для эукариот и бактерий является наличие в генах некодирующих участков – интронов, которых нет у бактерий. Причем структура РНК-полимеразы, компонентов транскрипционного комплекса, а также все дальнейшие процессы транскрипции и дальнейшей обработки (процессинга) мРНК у эукариот и архей очень схожи, в то время, как у бактерий существенно отличаются. Например, транскрипция и трансляция, на матрице синтезируемой мРНК, у бактерий идут одновременно и для старта синтеза белка не требуется не требуется процессинга мРНК. Причем, трансляция бактерий начинается не с метионина, как у эукариот (и архей), а с формилметионина.
Помимо особенностей, связанных с транскрипцией и трансляцией, для прокариот, в отличие от эукариот, характерно большое разнообразие метаболических особенностей, таких как способность к метаногенезу архей, хемолитоавтотрофность, способность к фиксации азота и способность к аноксигенному фотосинтезу.
Исходя из этого, становится видно, что все три выделенные на настоящий момент домена – бактерии, археи и эукариоты существенно отличаются друг от друга. Причем археи, хоть и являются прокариотами и несут в своем строении типичные прокариотические черты – отсутствие ядра и мембранных органоидов в цитоплазме, кольцевая ДНК, кольцевая хромосома и многое другое, тем не менее в некоторых чертах похожи на эукариот. Говоря о родстве между этими тремя группами, стоит отметить, что согласно доминирующей в настоящее время гипотезе, считается, что не смотря на то, что и бактерии, и археи относятся к прокариотам, последние все же более близки к эукариотам. Таким образом, в ходе эволюции сперва произошло разделение на группу бактерий и некого общего предка, от которого в дальнейшем произошли археи и эукариоты
В современной науке принято использовать термин «микробиота»
S – константа седиментации. Скорость осаждения частицы при ультрацентрифугировании. В данном контексте ее используют, чтобы охарактеризовать размер частицы.
# Микробиология
# 11 класс
Оболочка клеток прокариот
Клеточная стенка бактерий хорошо сохраняет форму клетки, т.к. это довольно прочная структура. Прочность оболочек связана с тем, что в них находятся муреины. Это молекулы, состоящие из параллельных полисахаридных цепей, перекрестно связанных короткими цепями аминокислот через регулярные промежутки. Получается своеобразный сетчатый каркас, окружающий клетку. На самом деле это одна огромная молекула.
Клеточная стенка предохраняет клетку от разрыва при поступлении в нее большого количества воды. Все необходимые вещества попадают в клетку через мельчайшие поры в клеточной стенке.
С клеточными стенками связан еще один метод классификации бактерий на группы: грамположительные и грамотрицательные.
Название групп связано с разработкой метода окрашивания бактерий. Изобретение метода принадлежит Кристиану Граму – датский биолог. Из-за различия в строении клеточной стенки, некоторые бактерии окрашиваются (грамположительные), а некоторые – нет (грамотрицательные).
У грамположительных бактерий в муреиновую сетку в основном встроены полисахариды и белки. Это делает стенку клетки более толстой.
Стенки грамотрицательных бактерий тоньше, но структура их более сложная. Муреиновый слой этих клеток покрыт тонким и гладким слоем, который состоит из липидов и полисахаридов. Они выполняют функцию дополнительной защиты для клетки. Он, например, выдерживает воздействие такого антибактериального фермента как лизоцим, который содержится в слезах или слюне.
Наличие этого слоя делает грамотрицательные бактерии устойчивыми против пенициллина. Ведь применение именно этого препарата спасло многие жизни от бактериальных инфекций.
Половой процесс солелюбивой археи Haloferax — нечто промежуточное между типичным прокариотическим и эукариотическим сексом
Если эукариотический секс — исходная черта эукариот, то имеет смысл поискать среди современных архей варианты генетического обмена, переходные между типичным прокариотическим и эукариотическим сексом. Крайне интересно было бы выяснить, как меняются генами локиархеи, но об этом пока ничего не известно. На сегодняшний день подробно изучена система генетического обмена только у одной археи — солелюбивого (галофильного) микроба Haloferax volcanii, обитателя Мертвого моря. Об этом микробе рассказано в новости Альтернативный способ репликации ДНК оказался эффективнее традиционного. У Haloferax обнаружена система передачи генетического материала, действительно похожая на что-то промежуточное между обычным прокариотическим ГПГ (например, конъюгацией бактерий) и эукариотическим сексом (рис. 3).
Клетки Haloferax спариваются, соединяясь цитоплазматическими мостиками, по которым геномная ДНК может передаваться в обе стороны, то есть обе клетки могут быть как донорами, так и реципиентами генетического материала. При этом могут передаваться крупные фрагменты хромосомы (длиной до 13% генома как минимум). В лаборатории можно добиться даже слияния клеток: для этого нужно понизить концентрацию Mg2+ в среде, что приводит к растворению клеточных стенок и дестабилизации цитоплазматических мостиков (рис. 3, справа). Предполагается, что такое слияние иногда может происходить и в природе.
В 2010 году была предложена красивая гипотеза, связывающая воедино происхождение эукариот и полового размножения и опирающаяся в том числе на данные по половому процессу Haloferax (см.: J. Gross, D. Bhattacharya, 2010. Uniting sex and eukaryote origins in an emerging oxygenic world, статья в открытом доступе). В статье обосновываются три положения:





1) Первые эукариоты появились в раннепротерозойских мелководных местообитаниях, когда концентрация кислорода начала расти, но озонового экрана еще не было. Под действием ультрафиолета на мелководьях должна была повыситься концентрация активных форм кислорода — сильных мутагенов. Появление эукариот стало закономерным итогом попыток архей защититься от этой напасти.
2) Архейные предки эукариот обменивались генами примерно так же, как современный Haloferax. Авторы выстроили гипотетический сценарий постепенного эволюционного перехода от полового процесса Haloferax к полноценному эукариотическому сексу.
3) Совершенствование системы генетического обмена было необходимо предкам эукариот, потому что они использовали чужую ДНК в качестве матрицы для починки разрывов двойной спирали ДНК при помощи гомологичной рекомбинации.
При всех достоинствах этой гипотезы, она не учитывает одну важную деталь, которая не противоречит первым двум пунктам, но сильно бьет по третьему. Дело в том, что Haloferax, как и многие другие археи, является полиплоидом. В каждой клетке Haloferax содержится в среднем по 17 копий генома (кольцевых хромосом).
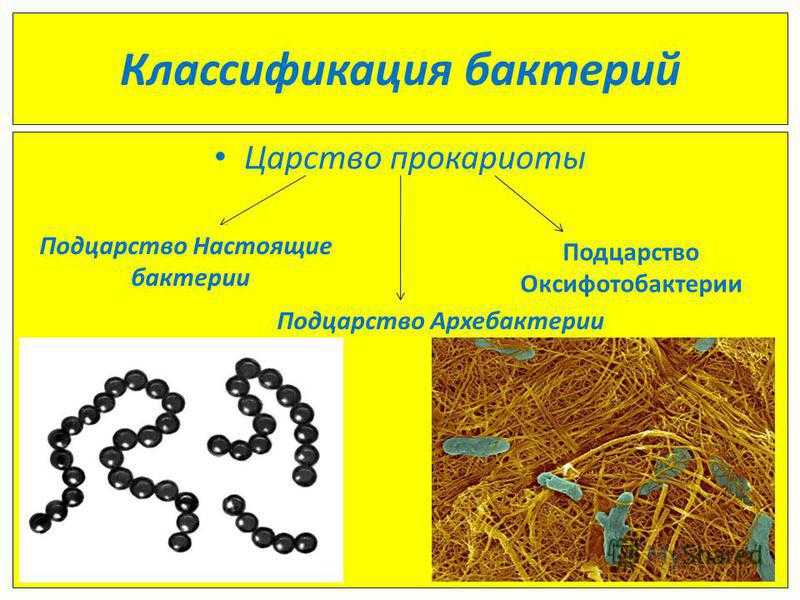
Представители царства прокариотов
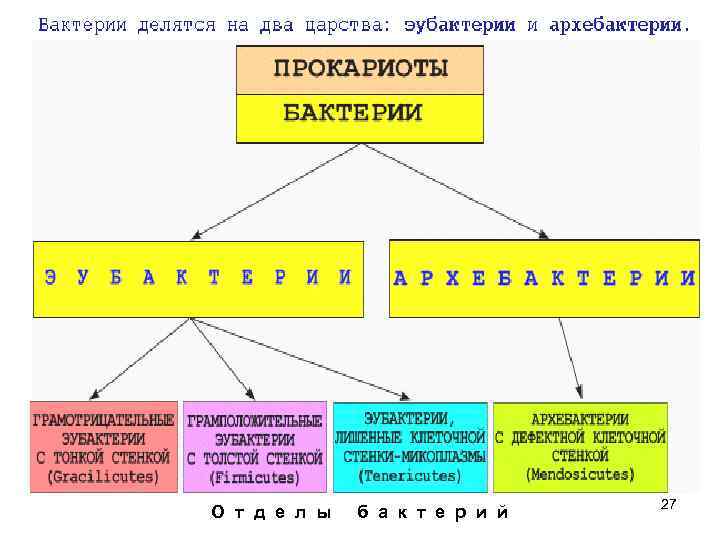
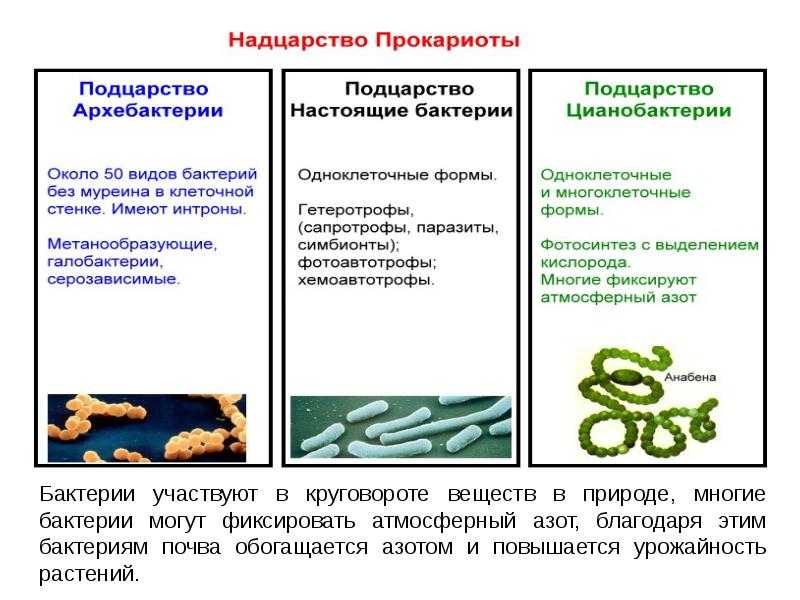
Название «прокариоты» происходит от древнегреческих слов «перед» и «ядро», т. е. это организмы, существовавшие еще до появления в клетках ядер. Это своеобразные предки эукариотов – видов, которые имеют оформленное клеточное ядро.
Прокариоты – это одноклеточные бактерии, в которых нет четко оформленного ядра клетки, ограниченного ядерной оболочкой, и дополнительных мембранных органоидов. Вместо этого прокариоты используют структуру, состоящую из ДНК (дезоксирибонуклеиновой кислоты), белков и РНК (рибонуклеиновой кислоты).
Исключение составляют фотосинтезирующие организмы, например, цианобактерии (сине-зеленые водоросли), которые имеют плоские цистерны – мембранная структура, обеспечивающая процесс жизнедеятельности клетки. Эта группа бактерий содержит зеленые и синие пигменты и участвует в процессе фотосинтеза, насыщая атмосферу кислородом.
Еще один представитель царства прокариотов – археи (лат. «древний, первозданный»). Эти одноклеточные бактерии интересны не только тем, что у них нет ядра, но и особенностями питания. Так, они могут существовать и находить пищу в самых экстремальных условиях – в горячих источниках и соленых озерах. Археи широко распространены в океанах, почве, болотах, даже в организме человека. Эти бактерии играют одну из ведущих ролей в круговороте углерода и азота на нашей планете.
Итак, к прокариотам относятся все бактерии, включая сине-зеленые водоросли и археи. Некоторые ученые относят к царству прокариотов и вирусы, но общего мнения на их счет пока не существует, так как некоторые биологи не считают вирусы живыми организмами.
Считается, что прокариоты появились около 3,5 млрд лет назад, и только через 2,5 млрд лет после них возникли первые эукариотные клетки. Все микроорганизмы, входящие в царство прокариотов, так или иначе способствовали созданию и поддержанию жизни на Земле.
Размеры
Однако были описаны нанобактерии, имеющие размеры меньше «допустимых» и сильно отличающиеся от обычных бактерий. Они, в отличие от вирусов, способны к самостоятельному росту и размножению (чрезвычайно медленным). Они пока мало изучены, живая их природа ставится под сомнение.
При линейном увеличении радиуса клетки её поверхность возрастает пропорционально квадрату радиуса, а объём — пропорционально кубу, поэтому у мелких организмов отношение поверхности к объёму выше, чем у более крупных, что означает для первых более активный обмен веществ с окружающей средой. Метаболическая активность, измеренная по разным показателям, на единицу биомассы у мелких форм выше, чем у крупных. Поэтому небольшие даже для микроорганизмов размеры дают бактериям и археям преимущества в скорости роста и размножения по сравнению с более сложноорганизованными эукариотами и определяют их важную экологическую роль.
Происхождение, эволюция, место в развитии жизни на Земле
Эукариоты возникли в результате симбиогенеза из бактериальных клеток намного позже: около 1,9—1,3 млрд лет назад. Для эволюции бактерий характерен ярко выраженный физиолого-биохимический уклон: при относительной бедности жизненных форм и примитивном строении, они освоили практически все известные сейчас биохимические процессы. Прокариотная биосфера имела уже все существующие сейчас пути трансформации вещества. Эукариоты, внедрившись в неё, изменили лишь количественные аспекты их функционирования, но не качественные, на многих этапах циклов элементов бактерии по-прежнему сохраняют монопольное положение.
Одними из древнейших бактерий являются цианобактерии. В породах, образованных 3,5 млрд лет назад, обнаружены продукты их жизнедеятельности — строматолиты, бесспорные свидетельства существования цианобактерий относятся ко времени 2,2—2,0 млрд лет назад. Благодаря ним в атмосфере начал накапливаться кислород, который 2 млрд лет назад достиг концентраций, достаточных для начала аэробного дыхания. К этому времени относятся образования, свойственные облигатно аэробной Metallogenium.
Появление кислорода в атмосфере нанесло серьёзный удар по анаэробным бактериям. Они либо вымирают, либо уходят в локально сохранившиеся бескислородные зоны. Общее видовое разнообразие бактерий в это время сокращается.
Предполагается, что из-за отсутствия полового процесса, эволюция бактерий идёт по совершенно иному механизму, нежели у эукариот. Постоянный горизонтальный перенос генов приводит к неоднозначностям в картине эволюционных связей, эволюция протекает крайне медленно (а, возможно, с появлением эукариот и вовсе прекратилась), зато в изменяющихся условиях происходит быстрое перераспределение генов между клетками при неизменном общем генетическом пуле.



